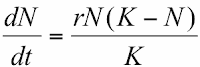What does it mean?
Assume that a predator feeds on two distinct and randomly distributed prey types (types 1 and 2 with densities N1 and N2, respectively). It is also assumed that the forager is attempting to maximize the rate by which energy is acquired. The predator searches the environment randomly (with search rate a) and encounters the two prey types in proportion to their respective densities. Let also the prey types differ in energetic reward (ei) and the time it takes to handle each prey after it has been captured and until ingested (handling time - hi). The two prey type differ in profitability such that e1/h1 > e2/h2. During a foraging bout when the predator does nothing but searching for and handling prey, the optimal strategy - the strategy that maximizes energy intake rate - is to always attack prey type 1 whenever encountered. Prey type 2 should always be left unattacked unless the density of prey type 1 falls below a certain threshold, namely the one given in the diet choice inequality above. If this criterion is satisfied both prey types should be attacked whenever encountered.
The left-hand side increases with increasing energetic reward from type 2 (e2) and decreasing handling time (h2). That is, the more profitable prey type 2 is, the higher the density threshold is.
The diet choice model implies an all-or-nothing response (the "zero-one rule")- either the less profitable prey type should be fully included in the diet or it should not be eaten at all.
The diet should be broad if the profitabilities are more similar and if the more profitable prey is relatively rare. A narrow diet would be advantageous if the converse is true.
For models (and experimental results) with less abrupt switches between the inclusion of both prey types or not, see for example Fryxell & Lundberg (1997).
Where does it come from?
In 1966 John Emlen and Robert MacArthur together with Eric Pianka laid the foundation for a new discipline subsequently christened "behavioral ecology" (MacArthur & Pianka 1966, Emlen 1966). They did so by assuming that organisms should one way or another behave optimally as a result of natural selection in order to maximize Darwinian fitness. A large number of optimality models emerged from from this then rapidly growing research field and the diet choice model was one of the first (see Stephens & Krebs 1986 for review).
Importance
The diet choice model together with the marginal value theorem launched the research program called "optimal foraging theory" and a huge number of models of foraging behavior with different currencies to maximize (other than energy gain), trade-offs, risk-sensitivity, foraging modes, and implications for population and community dynamics (Fryxell & Lundberg 1997). The diet choice model is closely related to the Disc equation and therefore an important link between fitness maximization at the individual level and the dynamics of predators and their prey.
The foraging decision based on the diet choice model is a response to the threat of lost opportunities. A predator that always attacks both prey indiscriminately would waste the opportunities for feeding on the more profitable prey.
Per Lundberg
Literature
Emlen, J.M. 1966. The role of time and energy in food preference. Am. Nat. 100: 611-617.
Fryxell, J. M. & Lundberg, P. 1997. Individual behavior and community dynamics. Chapman & Hall, NY.
MacArthur, R. M. & Pianka, E.R. 1966. On optimal use of a patchy environment. Am. Nat. 100: 603-609
Stephens, D. W. & Krebs, J. R. 1986. Foraging theory. Princeton UP.
Stephens, D. W., Brown, J. S. & Ydenberg, R. C. (eds) 2007. Foraging - behavior and ecology. Chicago UP